Заразный рак
Зара́зный рак (англ. transmissible cancer), или трансмисси́вная злока́чественная о́пухоль — форма рака, при которой сами раковые клетки или их скопления могут передаваться от одной особи к другой, при этом у особи-реципиента они не отторгаются иммунной системой, а приводят к развитию рака. Иными словами, в случае заразного рака инфекционным агентом являются сами раковые клетки. Достоверно известны три случая заразного рака: трансмиссивная венерическая опухоль собак, лицевая опухоль тасманийского дьявола и трансмиссивная лейкемия двустворчатых моллюсков. У человека на данный момент не известно заразных форм рака.
Трансмиссивная венерическая опухоль собак
[править | править код]Первым описанным случаем заразного рака стала трансмиссивная венерическая опухоль собак (англ. canine transmissible venereal tumor, CTVT). Помимо собак, заболевание обнаружено у волков, койотов и шакалов. Возможность передачи этой опухоли от одной особи к другой экспериментально продемонстрировали в 1876 году. Идентичность клеток CTVT стала очевидной после выявления одинаковых хромосомных перестроек, а также обнаружения интеграции ретротранспозона LINE1[англ.] перед геном Myc у всех клеток опухоли. Передача опухоли (плотного образования до 10 см в диаметре) происходит при половом контакте, и у новой особи опухоль, как правило, развивается на гениталиях. Любопытно, что клетки CTVT постоянно заимствуют митохондриальные геномы своих хозяев. Вероятно, митохондриальные геномы опухолевых клеток быстро приходят в негодность из-за многочисленных мутаций. Как правило, CTVT в течение нескольких месяцев после активного роста регрессирует, не приводя к гибели собаки, однако в некоторых случаях регрессии опухоли не происходит. Описаны даже случаи метастазирования CTVT. Орган или ткань, от которых произошли клетки CTVT, неизвестны[1][2][3].
По оценкам, CTVT появилась очень давно — от 10 тысяч до 12 тысяч лет назад, что делает ее самой древней самоподдерживающейся линией клеток[англ.] (первой искусственной самоподдерживающейся линией стала HeLa, произошедшая от клеток рака шейки матки). Она обнаружена у собак на всех континентах, кроме Антарктиды[2]. Предполагают, что CTVT привела к гибели древнейших американских домашних собак, прибывших на континент вместе с первыми поселенцами около 15 тысяч лет назад. Судя по анализу древних геномов, клетки CTVT генетически гораздо ближе к первым американским собакам, чем к современным животным, из-за чего иммунная система первых не была в состоянии эффективно отторгать опухоль[4].
Лицевая опухоль тасманийского дьявола
[править | править код]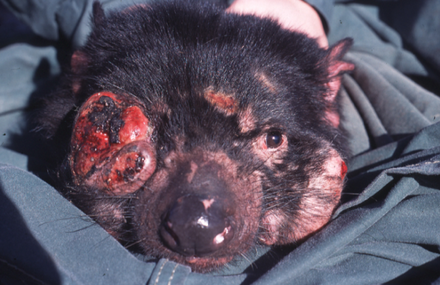
Впервые лицевая опухоль тасманийского дьявола (англ. devil facial tumor disease 1, DFTD1) была описана в 1996 году на северо-востоке Тасмании. Передача этой крайне агрессивной опухоли происходит при укусах, которые служат важным средством социального общения животных. Трансмиссивный характер опухоли стал очевиден после обнаружения уникальных хромосомных перестроек во всех опухолевых клетках, а также после полного секвенирования геномов[англ.] клеток двух различных опухолей[5].
В 2014 году на юго-востоке Тасмании обнаружили другую линию раковых клеток лицевых опухолей тасманийского дьявола (DFTD2). Пока выявлено 5 особей с DFTD2, причем все они были самцами. Родоначальником DFTD1 была самка, в то время как DFTD2 появилась у самца. Возможно, самки каким-то образом распознают клетки DFTD2 как чужеродные и избавляются от них, однако делать какие-либо заключения преждевременно в силу недостатка данных. Эпидемиология DFTD2 еще не изучена, но проявления DFTD1 и DFTD2 очень схожи: оба заболевания сопровождаются ростом опухолей до 10 см в диаметре на лице и шее животных. Со временем опухоли подвергаются некрозу и воспаляются, приводя к значительным изменениям анатомии челюсти животного, из-за которых оно не может питаться и пить. Кроме того, в 65 % случаев лицевая опухоль метастазирует[1][2].
Генетические маркеры и кариотипы указывают, что DFTD1 и DFTD2 возникли у разных особей. Клетки DFTD1 произошли от шванновских клеток, образующих миелиновую оболочку периферических нервов, а происхождение клеток DFTD2 еще неизвестно. В отличие от опухоли собак, DFTD всегда приводит к летальному исходу, что представляет серьёзную угрозу для тасманийского дьявола как вида. Описаны только 6 животных, у которых развился иммунный ответ на клетки опухоли[1][2][3].
Трансмиссивная лейкемия двустворчатых моллюсков
[править | править код]По меньшей мере 15 видов двустворчатых моллюсков подвержены смертельной трансмиссивной лейкемии. Опухолевые клетки при этом заболевании происходят от гемоцитов — клеток, циркулирующих в гемолимфе. Первые случаи заболевания описаны в 1970-х годах, и с тех пор болезнь начала быстро распространяться по восточному побережью Северной Америки. Иногда происходят вспышки заболевания, когда более 90 % популяции оказывается зараженной. Клетки разных видов трансмиссивной лейкемии характеризуются значительным увеличением числа копий ретротранспозонов в одних и тех же участках генома[1][2][3].
Возможно, перемещение ретроэлементов запускается факторами внешней среды, такими как перенаселенность, загрязнение, изменение температуры воды. К настоящему времени трансмиссивная лейкемия детально изучена у песчаной мии (Mya arenaria), мидий Mytilus trossulus[англ.], съедобной сердцевидки (Cerastoderma edule) и Polititapes aureus, и у каждого вида имеется своя независимая линия трансмиссивной лейкемии. У сердцевидки обнаружены две независимые трансмиссивные лейкемии, как у тасманийского дьявола[1][2][3].
Интересно, что лейкемия Polititapes aureus была получена им от другого вида двустворчатых моллюсков — Venerupis corrugata[англ.], — который в настоящее время лейкемии не подвержен, несмотря на общность местообитания с P. aureus. Вероятно, ему удалось приобрести резистентность к этому заболеванию. Передача раковых клеток происходит через морскую воду, которая проникает в организм моллюска при фильтрации. Со временем раковые клетки распространяются по всем органам и тканям заражённого животного[1][2][3].
Заразный рак у человека
[править | править код]Передача раковых клеток от одного человека к другому — чрезвычайно редкое явление. Как правило, она происходит при пересадке органов, так как иммунная система реципиента искусственно угнетается с целью предотвратить отторжение трансплантата. Другие случаи относятся к передаче раковых клеток от матери плоду, перемещению лейкемийных клеток между близнецами в утробе матери, а также передаче клеток внеплодной хориокарциномы от плода матери[6]. Недавно описали заражение пациента со СПИДом раковыми клетками обитающего в его кишечнике карликового цепня. Во всех этих случаях целостность физических и иммунных барьеров организма была нарушена[1][2].
Известно всего лишь несколько случаев передачи чужеродных раковых клеток людям с нормальным иммунитетом. Один хирург случайно занес себе в руку раковые клетки во время операции. Другой случай произошел с исследователем, который по неосторожности занес себе иглой клетки из линии аденокарциномы, что привело к образованию небольшой опухоли. В этих двух случаях опухоли успешно удалили без дальнейших рецидивов[2].
Иммунология заразного рака
[править | править код]Клетки любой раковой опухоли, как заразной, так и незаразной, вынуждены избегать действия иммунной системы хозяина, чтобы спастись. Один из современных способов лечения рака основан на том, чтобы вернуть раковые клетки под надзор иммунитета. Перед клетками заразного рака стоит еще более трудная задача. Чтобы заразить другую особь, им необходимо каким-то образом спастись от иммунной системы, для которой они являются чужеродными клетками и, следовательно, подлежат уничтожению. У челюстноротых позвоночных животных ключевую роль в распознавании своего и чужого играют молекулы главного комплекса гистосовместимости (англ. main histocompatibility complex, MHC). В клетках CTVT наблюдается пониженная экспрессия генов MHC, а также генов, связанных с презентацией антигенов и апоптозом[1][2].
Предполагают, что тасманийские дьяволы не развивают эффективного иммунного ответа против опухоли в силу низкого разнообразия MHC. Дело в том, что в начале XX века численность популяции дьяволов резко сократилась, что привело к снижению генетического разнообразия. Впрочем, в экспериментах по трансплантации кожи одного дьявола другому отторжение трансплантата всё-таки происходило. Оказалось также, что клетки DFTD тоже понижают экспрессию MHC, как и клетки опухоли собак. У людей перенос раковых клеток от матери к плоду возможен, потому что эмбрион имеет один аллель MHC от матери, и, если опухолевые клетки потеряют тот аллель, которого у эмбриона нет, они не будут распознаны плодом как чужеродные. У двустворчатых моллюсков, как и у всех беспозвоночных, MHC нет. Однако какой-то механизм распознавания своего и чужого у них всё-таки существует, так как их лейкемии, как правило, связаны с одним конкретным видом. Возможно, это достигается при помощи того же механизма, что препятствует слиянию колоний оболочников разных видов[1][2].
Стоит отметить, что клетки CTVT и DFTD сохраняют способность к экспрессии генов MHC, а не утрачивают ее необратимо. Например, если обработать клетки DFTD1 интерфероном γ, экспрессия генов MHC резко возрастет, а также восстановится способность к процессингу[англ.] и презентации антигенов. Вышеупомянутые шесть тасманийских дьяволов, развившие иммунный ответ против опухоли, имеют антитела к опухолевым клеткам, синтезирующим MHC, и у двух животных даже произошла спонтанная регрессия опухоли. У одного из дьяволов в опухоль проникли CD4+ и CD8+ T-лимфоциты. Неизвестно, сформировалась ли у этих животных иммунологическая память к опухолевым клеткам[1][2].
Регрессия CTVT, которая наблюдается у большинства собак, начинается с увеличения доли опухолевых клеток, экспрессирующих MHC. Вместе с этим CD4+ и CD8+ T-лимфоциты начинают проникать в опухоль и оказывают на ее клетки цитотоксическое действие. Вероятно, интерлейкин 6 и интерферон γ, вырабатываемые Т-лимфоцитами в опухоли, запускают образование MHC в раковых клетках[2].
Опухолевые клетки, не экспрессирующие MHC, могут стать жертвами других клеток иммунной системы — натуральных киллеров. Чтобы избежать этого, клетки CTVT выделяют цитокины, подавляющие иммунный ответ, такие как TGF-β. Секреция TGF-β наблюдается в фазе роста опухоли и на первых этапах стационарной фазы. Когда T-лимфоциты начинают синтезировать интерлейкин 6 и интерферон γ, развивается более мощная воспалительная реакция, нивелирующая эффект TGF-β. Рост DFTD, по всей видимости, от TGF-β не зависит[2].
Эволюция линий клеток заразного рака
[править | править код]Генетически клетки заразных опухолей, да и многих раковых опухолей вообще, отличаются от своих хозяев. Если жизнь «обычных» раковых клеток обрывается вместе со смертью хозяина, то время существования линии заразных раковых клеток существенно превышает время жизни одной особи за счет передачи от одного организма к другому. В случае CTVT линия заразных раковых клеток существует более 10 тысяч лет. Поскольку эволюция и размножение таких клеток совершенно независимы от хозяина, возникает соблазн даже рассматривать линии заразных раковых клеток как самостоятельные виды[1].
В принципе, процесс метастазирования, то есть смены местообитания раковых клеток в пределах одного организма, схож с передачей раковых клеток от одной особи к другой. В обоих случаях раковые клетки эволюционируют по Дарвину, приобретая мутации, которые потом отбраковываются естественным отбором сообразно условиям окружающей среды. Кроме того, и те, и другие клетки вынуждены изобретать разнообразные приспособления, чтобы избежать уничтожения иммунной системой хозяина[2].
Клетки CTVT характеризуются уникальной для опухолевых клеток стабильностью. В них почти не происходят мутации и хромосомные перестройки, и клетки в пределах опухолей удивительно однородны. На основании этого ученые заключили, что эволюция клеток CTVT, вначале сопровождавшаяся многочисленными мутациями и перестройками, в данный момент вышла на плато. Клетки CTVT достигли тонкого равновесия со своим хозяином: с одной стороны, они не убивают его, однако долгое время, вплоть до полной регрессии опухоли, собака-хозяин служит в роли переносчика опухолевых клеток. Клетки DFTD, напротив, находятся на ранних этапах эволюции и еще далеки от равновесия со своим хозяином[3].
Активация ретротранспозонов в клетках, ставших впоследствии клетками лейкемии двустворчатых моллюсков, вероятно, стала следствием чрезвычайной геномной нестабильности. Эти клетки содержат больше ДНК, чем нормальные клетки, и нередко являются анеуплоидными или тетраплоидными. Более того, клетки лейкемии в пределах одного организма могут различаться числом некоторых микросателлитов и заменами в митохондриальной ДНК[3].
SCANDAL (эволюционная гипотеза)
[править | править код]В начале 2019 года в статье нескольких исследователей[7] была сформулирована и проанализирована гипотеза о том, что, поскольку известные клетки заразного рака могут рассматриваться как одноклеточные паразиты, эволюционировавшие из многоклеточных организмов, то когда-то подобные клетки могли в итоге эволюционировать в многоклеточные организмы, чьи потомки дошли до наших дней. В работе был проведен анализ геномов четырех выбранных групп организмов — из типов Placozoa, Dicyemida, Orthonectida и подкласса Myxosporea. Для первых трех выбранных групп «гипотеза SCANDAL» не подтвердилась, для последнего (Myxosporea) она представляется правдоподобной, но для окончательного вывода мало данных.
Примечания
[править | править код]- ↑ 1 2 3 4 5 6 7 8 9 10 11 Metzger Michael J., Goff Stephen P. A Sixth Modality of Infectious Disease: Contagious Cancer from Devils to Clams and Beyond (англ.) // PLOS Pathogens. — 2016. — 27 October (vol. 12, no. 10). — P. e1005904. — ISSN 1553-7374. — doi:10.1371/journal.ppat.1005904.
- ↑ 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 Siddle H. V. Cancer as a contagious disease (англ.) // HLA. — 2017. — 15 February (vol. 89, no. 4). — P. 209—214. — ISSN 2059-2302. — doi:10.1111/tan.12980.
- ↑ 1 2 3 4 5 6 7 Ujvari Beata, Papenfuss Anthony T., Belov Katherine. Transmissible cancers in an evolutionary context (англ.) // BioEssays. — 2016. — July (vol. 38). — P. S14—S23. — ISSN 0265-9247. — doi:10.1002/bies.201670904.
- ↑ Ní Leathlobhair Máire, Perri Angela R., Irving-Pease Evan K., Witt Kelsey E., Linderholm Anna, Haile James, Lebrasseur Ophelie, Ameen Carly, Blick Jeffrey, Boyko Adam R., Brace Selina, Cortes Yahaira Nunes, Crockford Susan J., Devault Alison, Dimopoulos Evangelos A., Eldridge Morley, Enk Jacob, Gopalakrishnan Shyam, Gori Kevin, Grimes Vaughan, Guiry Eric, Hansen Anders J., Hulme-Beaman Ardern, Johnson John, Kitchen Andrew, Kasparov Aleksei K., Kwon Young-Mi, Nikolskiy Pavel A., Lope Carlos Peraza, Manin Aurélie, Martin Terrance, Meyer Michael, Myers Kelsey Noack, Omura Mark, Rouillard Jean-Marie, Pavlova Elena Y., Sciulli Paul, Sinding Mikkel-Holger S., Strakova Andrea, Ivanova Varvara V., Widga Christopher, Willerslev Eske, Pitulko Vladimir V., Barnes Ian, Gilbert M. Thomas P., Dobney Keith M., Malhi Ripan S., Murchison Elizabeth P., Larson Greger, Frantz Laurent A. F. The evolutionary history of dogs in the Americas (англ.) // Science. — 2018. — 5 July (vol. 361, no. 6397). — P. 81—85. — ISSN 0036-8075. — doi:10.1126/science.aao4776.
- ↑ Kreiss A., Tovar C., Obendorf D. L., Dun K., Woods G. M. A Murine Xenograft Model for a Transmissible Cancer in Tasmanian Devils (англ.) // Veterinary Pathology. — 2010. — 22 September (vol. 48, no. 2). — P. 475—481. — ISSN 0300-9858. — doi:10.1177/0300985810380398.
- ↑ Greaves Mel, Hughes William. Cancer cell transmission via the placenta (англ.) // Evolution, Medicine, and Public Health. — 2018. — 1 January (vol. 2018, no. 1). — P. 106—115. — ISSN 2050-6201. — doi:10.1093/emph/eoy011.
- ↑ Panchin A. Y., Aleoshin V. V., Panchin Y. V. From tumors to species: a SCANDAL hypothesis // Biology Direct, 2019, 14:3
Литература
[править | править код]- Елизавета Минина. Заразный рак: правило или исключение? Статья на Биомолекула.ру.
Эта статья входит в число добротных статей русскоязычного раздела Википедии. |